1. Conodonts in Vertebrate Evolution
Go back to Analysis: Research In Progress
Conodonts were first recognized in the 1830's by the Russian paleontologist
Christian Pander. The earliest conodont teeth appeared
during the Cambrian about 500 million years ago and remained widespread
throughout the whole 300 million years of the paleozoic. However,
the actual bodies of these animals contained no bones and left a p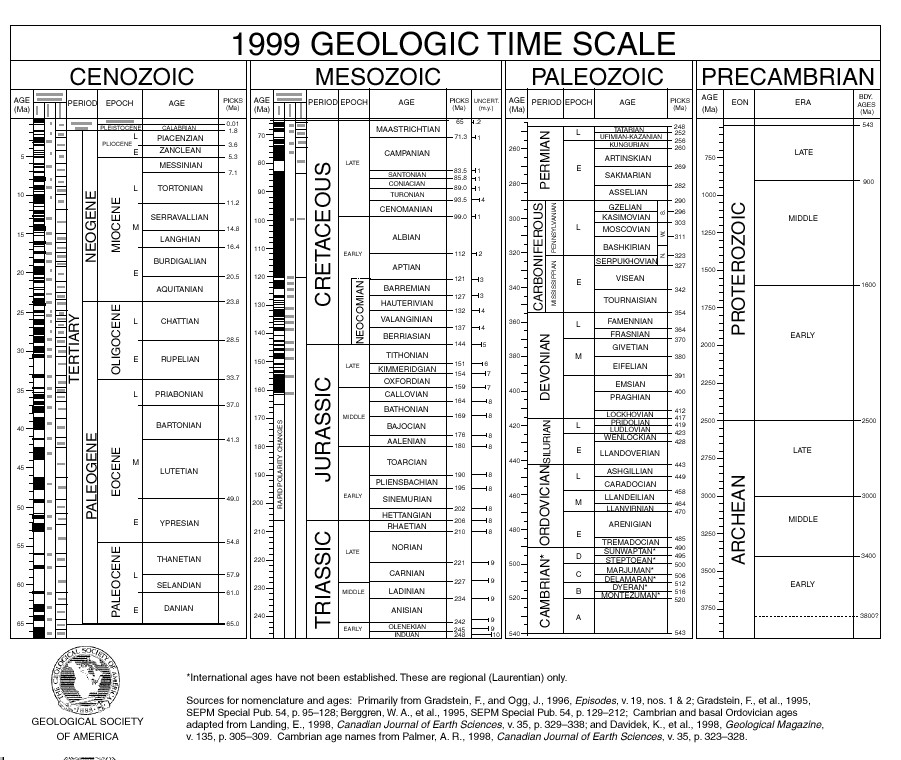 oor fossil
record. The first conodont body fossils were described by Briggs,
Clarkson and Aldridge in 1963. These delicate impressions dating
from the Lower Carboniferous ( from roughly 340 MYA) record a sophisticated
body plan that includes eyes and an assemblage of tooth elements arranged
at one end, connected to a few centimeters of worm-like body that
bears distinctive chevron-shaped muscle markings -- placing them in the
phylum of chordates, and making conodonts candidates for the first
vertebrates.
oor fossil
record. The first conodont body fossils were described by Briggs,
Clarkson and Aldridge in 1963. These delicate impressions dating
from the Lower Carboniferous ( from roughly 340 MYA) record a sophisticated
body plan that includes eyes and an assemblage of tooth elements arranged
at one end, connected to a few centimeters of worm-like body that
bears distinctive chevron-shaped muscle markings -- placing them in the
phylum of chordates, and making conodonts candidates for the first
vertebrates.
Until recently, the classic line of vertebrate evolution was thought to
start with "Pikaia"
-- a bug-sized
creature found in the 505-million year old Middle Cambrian Burgess Shale. Fossils of
this creature show what appears to be a longitudinal notochord flanked by
chevron-shaped structures that resemble muscle blocks. However, a new
convicing candidate for first chordate was announced in 1999 with Haikouichys --
an early (530 MYA) Cambrian fossil found in China. These 2 to 3-
cm fossils resemble a tiny fish -- the first such animal in the fossil
record. Better specimens were announced in 2003 which show well-developed
eyes, and other sensory structures characteristic of the cratiates, as well
as the muscle blocks typical of early vertebrates (Nature 421, pp 526-529).
Unmistakable fish fossils don't appear until the late Ordovician (450 MYA),
with the plate-covered 18-inch Bolivian fossils such as Sacabambaspis.
This jawless fish had paired gill structures and well defined anatomy,
including a brain. Animals with true jaws with teeth didn't arrive
until the late Silurian about 420 MYA with the first jawed fish (gnathostomes)
demonstrated in Acanthodes -- where the first two bony gill arches are
eliminated, and the third gill arch appears to have become modified into
a lower jaw, complete with teeth. By the early Devonian era -- the
"Age of Fishes" -- three other type of gnathostomes also appear --
the short-lived Placoderms, cartilagenous fish (such as sharks, which
contain no cellular bone), and more familiar bony fish that survive today
as the Telosts. During this entire vast span of paleozoic time,
conodonts were amazingly successful and diversified many times, waxing and
waning in genus diversity. By the time codononts died out in the Triassic,
the first Dinosaurs were roaming the continents. Although they may
represent an evolutionary sideline, conodonts can still claim the distinction
of demonstrating nature's first experiments with teeth -- teeth that had
a surprisingly sophistocated architecture.
This observation raises an interesting puzzle. Bone was
classically considered to be derived from the primitive, plate-like dermal
scales on the first jawed fish that appeared in the Silurian. More
recently (Science Vol 299, No 5610 pg 1235) regularly renewed teeth
were found on late placoderms (the arthrodires) suggesting that teeth appeared
twice in two widely seperate vertebrates by covergent evolution. However,
with their enamel, cellular bone-like structure and specialized differentiated
denticles, conodont elements also have a surprisingly modern, almost
mammel-like appearance from an evolutionary standpoint.
Yet conodonts first appear in the late Cambrian -- about 50 million years
before the first appearance of jawless fish in the late Ordovician.
Did bony enamel teeth evolve three times -- or is classical thinking mistaken?
Could conodonts be a "missing link" in our own fossil history?
Perhaps some feature of conodont teeth prevented them from growing to large
size, forcing early fish to develop a secondary specialization, substituting
dermal plates for teeth. If this is true, much of the actual work
of evolutionary change might have taken place in these soft-bodied creatures
out of view of the fossil record. However this is conjecture,
since, no conodont with true jaws has ever been found.
Although the conodont feeding apparatus contains advanced cellular bone
with differentiated teeth, the mouth is utterly different from that
of the jawed fish. Conodonts possessed up to 7 very different
types of bony elements working mysteriously together. Although many
different varieties of conodonts have been described, the chewing apparatus
seemed to be a three-stage process, first involving grasping spine or
comb-like elements, then shearing or cutting elements, and finally grinding
or slicing elements that eventually developed a peculiar rocker-type arrangement.
Yet the tooth elements are made of cellular bone and enamel -- only
found in modern animals. In fact, individually differentiated
teeth only appear in the earliest progenitors of Mammals -- such as the
pelycosaurs that appear in the late Periman 295 MYA.
There remain many other fundumental difference between mammals and the
early amphibians that metamorphesed into reptiles. Even at the very
earliest stages of the embryo, mammal egg cells exibit a unique spiral
clevage pattern, and many aspects of the fossil skulls of these creatures
remain poorly correlated with amphibian ancestors, suggesting a very ancient
common ancestor -- one that might even have preceded the first tetrapod land
animals. A recent clue about connections of these animals appears
in the 1998 with Eucritta.
(An excellentl book on this subject was published by Jennifer Clack
in 2002 -- Gaining
Ground: the origin and evolution of tetrapods.)
Condonts are extinct, but some relatives may still be alive
today. The hagfish
-- or "slime eel" -- is one such fascinating vertebrate. (See Early
Vertebrates 1998, by Philippe Janvier, pg 44) The boneless deep-sea
animal Myxine glutinosa features a unique body plan with an odd
cartilaginous skull with a single presasal opening where water is sucked
in and pumped out through gills using a specialized organ called the velum.
The simple brain is protected in a fiberous sheath, receiving limited
input from nearly vestigial eyes, and a primitive organ of equilibrium.
The heart has a peculiar design in which the atium and ventricle are separated,
and blood is pumped by three additional assessory venous hearts. The
jawless mouth is not connected to the respiratory system, and opens sideways
to unsheath rows of teeth that evert inside-out to bite and then retract
into the mouth, pulling the food into the digestive tract. On the deep ocean floor, these eels feed
by tying their bodies into knots, giving them the leverage needed to bite
food off of carrion they find on the ocean bottom. When captured, this
animal defends itself by exuding an extraordinary volume of slimy gel.
This strange creature may be our best clue to imagining the conodonts that
dominated the seas of Earth for almost half the history of animal evolution.
Amphioxus is another surviving representative of early chordates. The lancelet is a 6-centimeter headless eel-like animal that lacks eyes, bones or jaws -- but does have a mouth with sensory cilia, a notochord, post-anal tail, muscle blocks derived from embryonic somites. Unlike worms or insects, Amphioxus is a deuterostome (meaning the early embryo digestive tract forms from the anus and tunnels forward toward the mouth.) Over the last 550 million years, the amphioxus genome has twice been doubled and kneeded like dough to produce the complex array of genes that govern modern vertebrates development -- including complex organs like the brain, kidney, pituitary and pancrease. DNA sequencing suggests that two genome-doubling events occurred after Amphioxus, but before the appearance of cartilaginous fish -- an era of evolutionary history that was dominated by the mysterious conodonts.
Note:
Rapid progress is being made today on the genetic origin of jaws. (See
perspective article Jaws of the fates by Koentges and Matsuoka, Science
vol 298 p 371 11 Oct 2002, and the article: Specification of jaw subdivisions
by Dix genes by Depew, Mighael Lufkin and Rubenstein, Science vol
298 p 381 11 Oct 2002) The embryo uses neural crest cells
to create granchial arches that are transformed by the action of three
nested pairs of homeobox genes Dix 1/2, Dix 5/6 and Dix3/7 which shape
the bones of the jaws. Differences in the jaw structure among the
kinds of vertebrates are caused by different behavior patterns of the
target cells. Jawless fish such as lampreys possess only two pairs
of Dix genes that are not nested.
There are additional lines of research that may eventually throw some
light on this matter. In Science (Vol 282 No 5391 p 1136) mammal tooth
differentiation was more directly connected with homeobox Msx-1 and Barx-1
genes. It would be interesting to see if these genes are present in
sharks, fish, and hagfish.
Page established 7-7-99, updated 9-24-2009 -- copyright Jim Davison
Go back to: Analysis: Research In Progress--
Please mail comments or suggestions to: My E-Mail Address